تار کشنده چیست؟
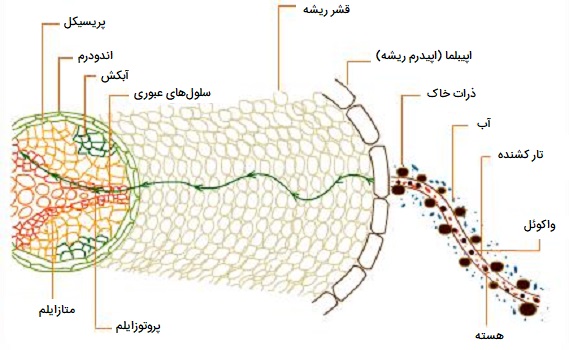
«تار کشنده» (Root hair) یا موهای ریشه که موهای جاذب نیز نامیده میشوند، رشد لولهای یک سلول اپیدرمی (سلول تشکیل دهنده مو در اپیدرم) ریشه هستند. این ساختارها امتداد جانبی یک سلول واحد هستند و به ندرت منشعب میشوند. آنها در ناحیه بلوغ (یا منطقه تمایز ریشه) یافت میشوند. درست قبل و در حین رشد سلولهای تار کشنده ریشه، فعالیت فسفوریلاز افزایش مییابد. گیاهان آب را از طریق ریشه از خاک با جریان تودهای جذب میکنند. سلولهای تار کشنده ریشه با افزایش سطح ریشه به منظور جذب آب بیشتر با این فرایند سازگار میشوند. واکوئل بزرگ در داخل سلولهای تار کشنده ریشه این مصرف را بسیار کارآمدتر میکند. سلولهای تار کشنده ریشه در نوک ریشههای گیاه رشد میکنند. سلولهای موی ریشه بین ۱۵ تا ۱۷ میکرومتر و طول آنها بین ۸۰ تا ۱۵۰۰ میکرومتر متفاوت است.

عملکرد ریشه
گیاهان برای اینکه قائم بمانند و تحت تأثیر باد قرار نگیرند به ریشه نیاز دارند. آب و مواد مغذی نیز از طریق سیستم ریشه وارد گیاه میشوند. تا زمانی که گیاه به طور کلی، از جمله داشتن شاخ و برگ، به رشد خود ادامه دهد، سیستم ریشه به کار خود ادامه میدهد. هنگامی که تعادل خاصی حاصل شد، گیاه به سادگی افراشتگی خود را حفظ کرده و رشد خود را متوقف میکند. تارهای کشنده ریشه جایی هستند که بیشتر مواد مغذی و آب جذب میشوند. نوک ریشه روزانه سلولهای جدیدی تولید میکند و در نتیجه موها نیز احتمال کنده شدن دارند. هنگامی که گیاه کمبود آب یا مواد مغذی دارد جذب بیشتری را برای تولید سلولهای بیشتر در نوک ریشه اختصاص میدهد. همچنین تارهای کشنده ریشه بیشتری تولید میشود تا زمانی که ریشه آن چیزی را که گیاه نیاز دارد (آب یا مواد مغذی بیشتر) پیدا کند. سپس قدیمی ترین تارهای کشنده ریشه از بین میروند.
تارهای کشنده ریشه فقط در ناحیه بلوغ یافت میشوند و در ناحیه طویل شدن وجود ندارند، احتمالاً به این دلیل که موهای ریشهای که به وجود میآیند با بلند شدن ریشه بریده شده و در خاک حرکت میکنند. تارهای کشنده ریشه به سرعت، حداقل ۱ میکرومتر در دقیقه رشد میکنند و این مسئله آنها را برای تحقیقات در مورد گسترش سلولها بسیار مفید میکند.

کار تار کشنده چیست؟
تارهای کشنده ریشه سطح مهمی را تشکیل میدهند زیرا برای جذب بیشتر آب و مواد مغذی مورد نیاز گیاه مورد نیاز هستند. آنها همچنین مستقیماً در تشکیل ندولهای ریشه در گیاهان حبوبات نقش دارند. ندولهای ریشه در ریشه گیاهانی مانند حبوبات یافت میشوند که با باکتریهای تثبیت کننده نیتروژن همزیستی ایجاد میکنند. تارهای کشنده ریشه در اطراف باکتری پیچ خورده و اجازه میدهند تا یک رشته عفونی از طریق سلولهای پوستی تقسیم شونده برای شکلگیری ندولها یا گرههای باکتریایی ایجاد شوند. در ندولها یا گرههای ریشه حبوبات، گاز نیتروژن (N۲) از اتمسفر به آمونیاک (NH۳) تبدیل میشود، که سپس توسط اسیدهای آمینه (بلوکهای سازنده پروتئینها)، نوکلئوتیدها (بلوکهای سازنده DNA و RNA) و همچنین مولکول مهم انرژی ATP و سایر اجزای سلولی مانند ویتامینها، فلاونها و هورمونها جذب میشود.
همچنین با داشتن سطح وسیعی کشیده شده در سلولهای تار کشنده، جذب فعال آب و مواد معدنی از طریق این ساختارهای ریشهای بسیار کارآمد است. سلولهای موی ریشه همچنین اسید (H+ را از اسید مالیک) ترشح میکنند که مواد معدنی را به شکل یونی تبادل کرده و حل کرده و جذب یونها را آسان میکند. هنگامی که یک سلول تار کشنده ریشه جدید رشد میکند، هورمونی را دفع میکند به طوری که سایر سلولها در مجاورت آن قادر به رشد تار کشنده نیستند. این امر توزیع برابر و کارآمد تارهای کشنده واقعی بر روی این سلولها را تضمین میکند و پوشش تارهای کشنده ریشه ثابت میماند.

تفاوت کرک و تار کشنده چیست؟
کرکها ساختارهای مو مانندی در برگ گیاهان هستند که در برخی از گونههای گیاهی وجود داشته و به حفظ دمای نرمال سطح برگ و گیاه و همچنین به تنظیم میزان تعرق و تبخیر آب از گیاه کمک میکنند اما همانطور که بیان شد تارهای کشنده سلولهای طویل شدهای در ریشه گیاه هستند که در جذب آب و مواد معدنی از خاک به گیاه نقش دارند.
عملکرد تار کشنده چگونه است؟
بیشتر جذب آب در تار کشنده ریشه اتفاق میافتد. موهای ریشه بلند هستند که به آنها اجازه میدهد تا بین ذرات خاک نفوذ کرده و از ورود موجودات باکتریایی مضر به داخل گیاه از طریق آوند زایلم (چوب) جلوگیری کنند. سطح مقطع سلول تار کشنده ریشه شکل تقریباً مستطیلی با دمی بلند و نازک که به سمت راست امتداد دارد و هستهای در بالا سمت چپ قرار دارد. این امر به این دلیل اتفاق میافتد که آب موجود در خاک دارای پتانسیل آبی بالاتری نسبت به سیتوپلاسم موهای ریشه است. عملکرد موهای ریشه جمع آوری آب و مواد معدنی (شیره خام) موجود در خاک و انتقال این محلول از طریق ریشه به بقیه گیاه است. از آنجا که سلولهای تار کشنده ریشه فتوسنتز نمیکنند، حاوی کلروپلاست نیز نیستند.
عملکرد تار کشنده ریشه برای تغذیه سالم گیاه ضروری است، اما عواملی در محیط وجود دارد که میتواند تغییر مثبتی در آنها ایجاد کند. بهبود ساختار تارهای کشنده ریشه که میتواند برای گیاه به دست آید بستگی به تعامل آنها با قارچهای اطراف دارد. قارچهای همزیست تولید کننده همزیستیهای میکوریز مانند میکوریزا آربوسکولار، ایجاد شده توسط قارچ های AM (میکوریز آربوسکولار (AM) نوعی قارچ میکوریزا است که در آن قارچ سیمبیون به سلولهای قشری ریشههای یک گیاه آوندی شکل میگیرد و به شکل آربوسکول در میآید) و ectomycorrhiza، ایجاد شده توسط قارچهای EM، با موهای ریشهای هستند که با آنها در تعامل هستند. به نظر میرسد این نوع میکوریزها به دلیل مزایایی که برای قارچ و گیاه دارند رایج هستند، میتوان آنها را در ۹۰ درصد از گونههای گیاهی زمینی طبقه بندی کرد.
شکل گیری این رابطه همزیستی برای قارچهای EM ابتدا با همزیستی با تار کشنده ریشه آغاز میشود. این فرایند زمانی شروع میشود که قارچ EM از خاک به تارهای کشنده ریشه میچسبند. قارچ سپس عوامل قابل نفوذی را ترشح میکند که تارهای کشنده ریشه به آنها بسیار حساس هستند و به این ترتیب اجازه میدهند که هیفها به داخل سلولهای اپیدرم نفوذ کرده و در اولین لایههای قشر ریشه یک شبکه «هارتیگ» (Hairtig) ایجاد کنند. این ساختار بسیار منشعب به عنوان رابط بین دو موجود زنده عمل میکند، زیرا سلولهای قارچی با تبادلاتی که به زودی بین گیاه و قارچ شکل میگیرند، سازگار میشوند. این فرایند شبیه به نحوه همزیستی قارچهای تار کشنده ریشه است، اما به جای عوامل قابل انتشار، آنها هیدرولازها را برای شل شدن دیواره سلولی ترشح میکنند که باعث ورود هیفهای میکوریز میشود و هیچ هارتیگ موجود نیست.

در تصویر بالا ریشهها و قارچهای میکوریز نشان داده شده است. ریشههای آربوسکولار – میکوریزی (AM)، به ویژه به وسیله تارهای کشنده ریشه، یونها را با محیط اطراف در تصویر A مبادله میکنند. ظرفیت آنها برای جذب مواد مغذی افزایش یافته است اما توسط قارچ AM جایگزین نمیشود. قارچهای اکتومیکوریزا (EM) اساساً با جلوگیری از تماس مستقیم بین خاک و نوک ریشههای کلونیزه شده (B و C) روابط گیاه و مواد معدنی را تغییر میدهند. نوک ریشه EM با میسلیوم پوشانده شده است و هیچ موی ریشه فعالی ندارد. از آنجایی که قارچهای EM تقریباً به تنهایی مسئول جذب مواد مغذی برای درخت میزبان هستند، طول هیف بسیار بیشتری نسبت به قارچهای AM دارند. دستههای طناب مانند هیفها (ریزومورفها) مواد مغذی را از دورترین هیفها به ریشهها منتقل میکنند.
تاثیر همزیستی قارچ ها با سلول های تار کشنده ریشه
تأثیرات زیادی از همزیستی قارچ در تارهای کشنده ریشه وجود دارد که ثابت میکند این رابطه برای گونههای گیاهی و قارچی مفید است، اما تأثیر اصلی گونههای قارچی در تارهای کشنده ریشه بر رشد آنها تمرکز میکند. مطالعات نشان داده است که قارچها در واقع رشد تارهای کشنده ریشه را بسته به کمبود آب یا مواد مغذی سازماندهی و کنترل میکنند. از آنجا که هر دوی این موجودات به مواد مغذی و آب احتیاج دارند، همکاری آنها با یکدیگر در صورتی ضروری است که هر دو بخواهند در محیط خود زنده بمانند. با تشخیص کمبود، واکنش تنش خشکی گیاه ایجاد شده و باعث رشد موهای ریشه میشود. قارچهای میکوریزا سپس از سیستم توسعه یافته خود برای کمک به گیاه در یافتن ناحیه مناسب تغذیه که نشان دهنده جهت رشد ریشهها است، استفاده میکنند.
کنترل جهت رشد موهای ریشه را میتوان به عنوان نقش مهمی در این رابطه همزیستی در نظر گرفت، زیرا در وقت و انرژی گیاه صرفه جویی شده و سعی میکند محل رشد آن را مشخص کند. این انرژی میتواند در عوض برای سایر فرآیندهای متابولیک گیاه مورد استفاده قرار گیرد که به نوبه خود به قارچ تغذیه کننده از این محصولات متابولیک کمک میکند.
شیره خام چیست؟
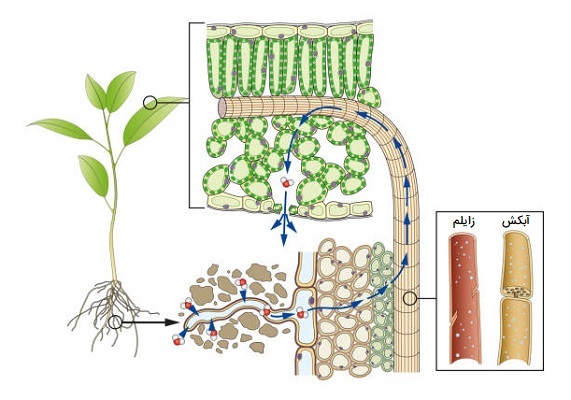
همانطور که یک انسان برای زنده ماندن، تغذیه میکند و نفس میکشد. یک گیاه نیز برای زنده ماندن، تغذیه و تنفس میکند. گیاه از طریق ریشههای خود و تارهای کشنده آب و مواد معدنی مورد نیاز خود را از خاک میگیرد اما باید این آب و نمکهای معدنی را تغییر دهد. برگ، آب و نمکهای معدنی را که از خاک توسط ریشهها گرفته شده است، تغییر میدهد. آب و نمکهای معدنی شیره خام را تشکیل میدهند. شیره خام یا شیره زایلم در درجه اول شامل یک محلول آبکی از هورمونها، عناصر معدنی و سایر مواد مغذی است. انتقال شیره خام در زایلم با حرکت از ریشه به سمت برگ مشخص میشود.
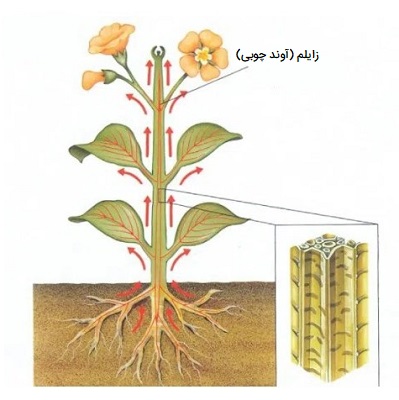
شیره خام چگونه در گیاه حرکت می کنند؟
زایلم یا آوند چوبی یک بافت ترکیبی در گیاهان آوندی است که به حمایت از گیاه کمک کرده و شیره خام را از ریشه به سمت بالا هدایت میکند. این ساختار از تراکئیدها، آوندها، سلولهای پارانشیم و الیاف چوب تشکیل شده است. ساختار زایلم به شکل لولهای بدون دیوارههای متقاطع است، که به ستون پیوسته آب اجازه میدهد و حمل و نقل سریعتر را در داخل لولهها تسهیل میکند. زایلم مسئول حرکت شیره خام به صورت یک جهته و همچنین مسئول جایگزینی آب از دست رفته در اثر تعریق و فتوسنتز است.
جذب شیره خام توسط تار کشنده ریشه
وقتی شیره خام از طریق اسمز وارد ریشهها میشود، سلولهای زایلم پر شده و متورم میشوند و به بیرونی ترین سلولهای سفت و سخت ریشه فشار وارد میکنند. این فشار، به ویژه هنگامی که سطوح آن در خارج از گیاه کم است، باعث میشود که شیره با وجود نیروی جاذبه مجبور به صعود به سمت بالا در گیاه شود. بار الکتریکی این سلولها از ریشه خارجی نوعی مسیر یک طرفه ایجاد میکند که اجازه نمیدهد شیره خام از ریشه به عقب و جلو برود. فشار ریشهای به عنوان فشاری بر روی عناصر آوندی زایلم در نتیجه فعالیتهای متابولیکی ریشه ایجاد شده است. فشار ریشهای یک فرآیند فعال است که با حقایق زیر تأیید میشود:
- سلولهای زنده در ریشه ضروری هستند تا فشار ریشه ایجاد شود.
- تامین اکسیژن و برخی از مهارکنندههای متابولیک بر فشار ریشهای بدون تأثیر بر نیمه تراوایی سیستمهای غشایی تأثیر میگذارد.
- مواد معدنی انباشته شده در برابر گرادیان غلظت با جذب فعال با استفاده از انرژی متابولیک تولید شده، پتانسیل آب سلولهای اطراف را کاهش داده و منجر به ورود شیره خام به سلولها میشود.
کشش تعرق مسئول صعود شیره خام در زایلم است. این افزایش شیره به عوامل فیزیکی زیر بستگی دارد:
- انسجام یا ویژگی هم چسبی مولکولهای آب. جاذبه متقابل بین مولکولهای آب یا شیره خام.
- کشش سطحی یا ویژگی دگر چسبی مولکولهای آب. مسئول بیشترین جاذبه بین مولکولهای آب یا شیره خام در فاز مایع است.
- چسبندگی. جذب مولکولهای آب یا شیره خام بر روی سطوح قطبی.
- فشار مویرگی. قابلیت بالا بردن شیره خام در لولههای یا آوندهای نازک.
شیره پرورده چیست؟
ویژگی شیره خام این است که از ریشه وارد شده و از طریق زایلم به بقیه گیاه منتقل میشود. پس از فرآیند فتوسنتز، به شیرهای لطیف تبدیل میشود که در جهت مخالف توسط آوندهای آبکش از برگها به ریشهها و سایر قسمتهای گیاه منتقل میشود. برگها مسئول تغییر شیره خام به شیره پرورده هستند. شیره پرورده در آب فقیرتر و غنی از مواد مغذی به دلیل عملکرد کلروفیل هستند. شیره پرورده تولید شده در برگها به منظور تغذیه گیاه به سمت ریشه فرود میآید. اکثر حشرات تغذیه کننده شیره از شیره پرورده یا شیره آبکش تغذیه میکنند، اگرچه برخی از حشرات از مایعات شیره خام نیز تغذیه میکنند.
تفاوت های شیره خام و شیره پرورده چیست؟
تفاوت اصلی بین آنها این است که شیره خام ترکیبی از نمکهای معدنی و سایر مواد است، در حالی که شیره پرورده از گلوکز، آب و مواد معدنی فتوسنتزی تشکیل شده است. شیره خام توسط آوند چوبی حمل میشود که از مواد محلول مانند آب، عناصر معدنی و تنظیم کنندههای رشد تشکیل شده است. از طریق یک لوله محکم از ریشه به برگ حرکت میکند. این شیره به برگها منتقل میشود، جایی که توسط فتوسنتز به شیره پرورده تبدیل میشود. شیره پرورده از مبدأ تشکیل آن از آبکش به ریشه و برگها و ساقههای سبز رنگ منتقل میشود.
شیره پرورده از آب، قندها، اسیدهای آمینه، ویتامینها، اسیدهای آلی، مواد معدنی محلول و تنظیم کنندههای گیاهی تشکیل شده است. در این مورد، فرضیه جریان فشاری به عنوان یک تکنیک حمل و نقل برای شیره پرورده قابل دسترسی است. این شیره توسط آبکش حمل میشود که نوعی بافت آوندی گیاهی است و میتواند در هر دو جهت جریان یابد و مواد مغذی را به تمام مناطق مورد نیاز گیاه منتقل کند، خواه اندام فتوسنتزی باشد یا نباشد.

سیستم جذب آب در گیاهان
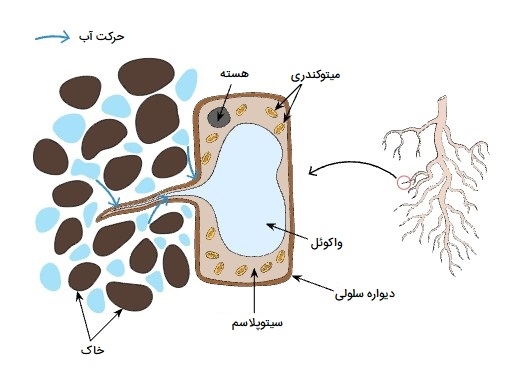
گیاهان این قابلیت را دارند که آب را از طریق تمام سطح خود از ریشه، ساقه، برگها، گلها و غیره جذب کنند. با این حال، از آنجایی که آب بیشتر در خاک موجود است، فقط سیستم ریشه زیرزمینی برای جذب آب تخصصی است. ریشهها اغلب گسترده هستند و به سرعت در خاک رشد میکنند. در ریشهها، موثرترین منطقه جذب آب، ناحیه تار کشنده ریشه است. آنها خروجیهای لولهای به طول ۵۰ تا ۱۵۰۰ میکرومتر (۰/۰۵ تا ۱/۵ میلیمتر) و ۱۰ نانومتر عرض هستند. هر تار کشنده ریشه دارای یک واکوئل مرکزی پر از شیره سلولی فعال اسمزی و یک سیتوپلاسم محیطی است. دیواره نازک و قابل نفوذ با مواد پکتیک در لایه بیرونی و سلولز در لایه داخلی است. تار کشنده ریشه به ریز منافذ مویرگی وارد میشوند، توسط ترکیبات پکتیک به ذرات خاک چسبیده و آب مویرگی را جذب میکنند.
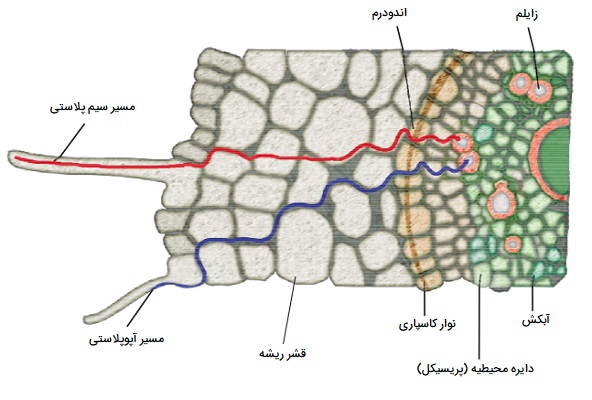
مسیرهای حرکت آب در ریشه
دو مسیر عبور آب از تارهای کشنده ریشه به آوند چوبی در داخل ریشه وجود دارد، این مسیرها آپوپلاستی و سیمپلاستی هستند که در ادامه در مورد هر کدام بیشتر توضیح داده ایم.
مسیر آپوپلاستی
در اینجا آب از تار کشنده ریشه به آوند چوبی از دیوارههای سلولهای مداخله گر بدون عبور از هیچ غشا یا سیتوپلاسم عبور میکند. این مسیر کمترین مقاومت را در برابر حرکت آب ایجاد میکند. با این حال، با وجود نوارهای کاسپاری غیرقابل نفوذ لیگنوسوبرین در دیواره سلولهای اندودرمی این مسیر انتقالی قطع میشود.
مسیر سیمپلاستی
آب از طریق پروتوپلاسم آنها از سلولی به سلول دیگر عبور میکند. آب به این طریق وارد واکوئلهای سلولی نمیشود. سیتوپلاسمهای سلولهای مجاور از طریق پلهایی به نام «پلاسمودِسماتا» (plasmodesmata) به هم متصل میشوند. برای ورود به مسیر سیمپلاستی، آب باید حداقل در یک مکان از پلاسمالما (غشای سلولی) عبور کند. به این مسیر عبور آب مسیر غشایی نیز میگویند. در حرکت سیمپلاستی با کمک جریان سیتوپلاسمی تک تک سلولها صورت میگیرد و کندتر از حرکات آپوپلاستی است. هر دو مسیر در حرکت در طول سراسر ریشه نقش دارند. آب از طریق آپوپلاست در قشر مغز جریان دارد. وارد مسیر سیمپلاست در اندودرم میشود که در آن دیوارها به دلیل وجود نوارهای کاسپاری در برابر جریان آب غیرقابل نفوذ هستند.
جذب آب میکوریزا
در میکوریزا تعداد زیادی هیف قارچی با ریشههای جوان همراه است. هیفهای قارچی با فاصله کافی در خاک گسترش مییابند. هیفها سطح وسیعی دارند و برای جذب آب و مواد معدنی تخصصی هستند. این دو به ریشه داده میشوند که هم قندها و هم ترکیبات حاوی نیتروژن را برای قارچ فراهم میکند. ارتباط قارچی بین ریشه و قارچ اغلب الزامی است. دانههای کاج و ارکیده جوانه نمیزنند و بدون تداعی میکوریزا به گیاه تبدیل میشوند.
مکانیسم جذب آب در گیاهان
جذب آب در ریشه بر دو نوع غیرفعال و فعال صورت میگیرد. در ادامه هر کدام از این مکانیسمها را جداگانه بررسی میکنیم.
جذب آب غیر فعال
نیروی جذب آب در این نوع مکانیسم از قسمتهای هوایی گیاه به دلیل از دست دادن آب در هنگام تعرق ایجاد میشود. این باعث ایجاد کشش یا پتانسیل کم آب چند اتمسفر در کانالهای آوند چوبی میشود. ایجاد کشش در کانالهای آوند چوبی گیاه به دلایل زیر صورت میگیرد:
- به دلیل فشار منفی که معمولاً در شیره خام زایلم یافت میشود، اگر برشی به شاخه داده شود آب بیرون نمیریزد.
- حتی در غیاب سیستم ریشه، آب میتواند توسط ساقه جذب شود.
- سرعت جذب آب تقریباً برابر با سرعت تعرق است.
تارهای کشنده ریشه به عنوان سیستمهای اسمزی کوچک عمل میکنند. هر تار کشنده ریشه دارای یک دیواره سلولی نفوذ پذیر نازک، یک سیتوپلاسم نیمه تراوا و یک شیره سلولی فعال اسمزی است که در واکوئل مرکزی وجود دارد. به دلیل دومی، یک سلول موی ریشه دارای پتانسیل آب ۳- تا ۸- بار است. پتانسیل آبی آب خاک ۰/۱ – تا ۰/۳ بار است. در نتیجه آب موجود در خاک وارد سلول تار کشنده ریشه میشود. با این حال، آب به واکوئل سلول نمیرود. در عوض به آپوپلاست و سمپلاست قشر مغز میرود، سلولهای اندودرمال و پریسیکل به دلیل پتانسیل بسیار کم آب ناشی از کشش که در آن آب وجود دارد و ناشی از تعرق در قسمتهای هوایی است، منفعلانه وارد کانالهای زایلم میشوند. یک گرادیان پتانسیل آب بین سلولهای تار کشنده ریشه، سلول قشر مغز، اندودرم، پریسیکل و کانالهای آوند چوبی وجود دارد به طوری که جریان آب قطع نمیشود.
جذب آب فعال
این جذب آب به دلیل نیروهای موجود در ریشه است. سلولهای زنده در شرایط متابولیک فعال برای این امر ضروری هستند. شناخته شده است که اکسینها جذب آب (حتی از محلول هایپرتونیک) را افزایش میدهند در حالی که مهارکنندههای تنفسی همان را کاهش میدهند. بنابراین انرژی (از تنفس) در جذب فعال آب نقش دارد. جذب آب از خاک و حرکت آن به داخل ممکن است در اثر اسمز اتفاق بیفتد. عبور آب از سلولهای زنده به مجاری زایلم میتواند به شرح زیر باشد:
- تجمع قندها یا نمکها در تراکئیدهای آوند چوبی به دلیل ترشح توسط سلولهای زنده مجاور و یا رها شدن در آنجا در طی پوسیدگی پروتوپلاستهای آنها.
- توسعه پتانسیل بیوالکتریک مطلوب برای حرکت آب به کانالهای زایلم.
- پمپاژ فعال آب توسط سلولهای زنده اطراف به تراکئیدها.
صعود شیره خام
شیره خام شامل آب و مواد محلول (مواد معدنی) است. حرکت آب به سمت بالا از ریشه به سمت نوک شاخههای ساقه و برگهای آنها صعود شیره نامیده میشود. از طریق عناصر آوندی آوند چوبی رخ میدهد. آب یا شیره از نزدیک نوک ریشه تا نوک ساقه در برابر نیروی جاذبه، گاهی تا ارتفاع ۱۰۰ متر بالا میرود. سرعت جابجایی ۲۵ تا ۷۵ سانتیمتر در دقیقه (۱۵ تا ۴۵ متر در ساعت) است. چندین نظریه برای توضیح مکانیسم صعود شیره ارائه شده است که در ادامه در مورد آنها توضیح داده ایم.
فرضیه نیروی حیاتی
نظریه نیروی حیاتی مشترک در مورد صعود شیره توسط J.C. Bose (1923) ارائه شد، به آن نظریه ضربان میگویند. این تئوری معتقد است که درونیترین سلولهای قشر ریشه آب را از سمت بیرونی جذب میکنند و همان را به کانالهای آوند چوبی پمپ میکنند. با این حال، به نظر نمیرسد که سلولهای زنده در صعود شیره نقش داشته باشند زیرا آب همچنان در گیاهی که ریشههای آن بریده شده است به سمت بالا میرود یا سلولهای زنده ساقه توسط سم و گرما کشته میشوند.
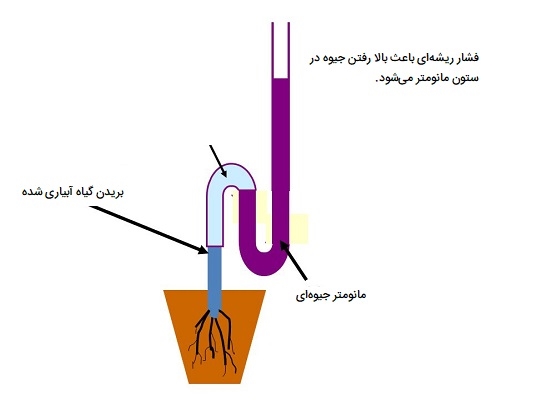
نظریه فشار ریشهای
این نظریه توسط پریستلی (۱۹۱۶) ارائه شد. فشار ریشهای فشار مثبتی است که در شیره آوند چوبی ریشه برخی از گیاهان ایجاد میشود. این تجلی جذب آب فعال است. فشار ریشهای در فصول خاصی مشاهده میشود که به نفع فعالیت متابولیک بهینه و کاهش تعرق است. این فشار در کشورهای گرمسیری در فصل بارندگی و در زیستگاههای معتدل در فصل بهار حداکثر است. مقدار فشار ریشهای معمولاً در گیاهان ۱ تا ۲ بار یا اتمسفر است. مقادیر بالاتر (به عنوان مثال، ۵ تا ۱۰ اتمسفر) نیز گاهی اوقات مشاهده میشود. فشار ریشهای در شرایط قحطی، دمای پایین، خشکسالی و کاهش دسترسی به اکسیژن، کاهش یافته یا از بین میرود. سه نقطه نظر در مورد مکانیسم ایجاد فشار ریشه وجود دارد که شامل موارد زیر هستند:
- فشار ریشهای اسمزی: تراکئیدهای اوند چوبی نمکها و قندها را جمع میکنند. غلظت زیاد املاح باعث خروج آب از سلولهای اطراف و همچنین از مسیر طبیعی جذب آب میشود. در نتیجه فشار مثبت در شیره زایلم ایجاد میشود.
- فشار ریشهای الکترواسموتیک: یک پتانسیل بیوالکتریک بین کانالهای زایلم و سلولهای اطراف وجود دارد که از عبور آب به داخل آنها جلوگیری میکند.
- فشار ریشهای غیر اسمزی: عناصر آوند چوبی متمایز کننده هورمونهایی تولید میکنند که به عنوان سینکهای متابولیک عمل میکنند و باعث حرکت آب به سمت آنها میشوند. سلولهای زنده اطراف زایلم میتوانند به طور فعال آب را به درون آنها پمپ کنند.
فشار ریشهای در همه گیاهان وجود ندارد. در بازدانگانی که دارای بلندترین درختان جهان هستند فشار ریشهای مشاهده نشده است. فشار ریشهای فقط در دورههای مطلوب رشد مانند بهار یا فصل بارانی مشاهده میشود. در این زمان شیره آوند چوبی نسبت به محلول خاک به شدت هایپرتونیک است و سرعت تعرق پایین است. در تابستان که نیاز به آب زیاد است، فشار ریشه به طور کلی وجود ندارد. فشار معمول مشاهده شده در ریشه معمولاً پایین است که نمیتواند شیره را به بالای درختان برساند.
حتی در غیاب ریشه آب همچنان به سمت بالا میرود. گیاهانی که به سرعت تعرق میکنند هیچ فشار ریشهای را نشان نمیدهند. در عوض فشار منفی در اکثر گیاهان مشاهده میشود. فشار ریشهای در شرایط نامساعد محیطی از بین میرود در حالی که صعود شیره بدون وقفه ادامه دارد. فشار ریشه معمولاً در شب هنگامی که تبخیر و تعرق کم است مشاهده میشود. ممکن است در برقراری مجدد زنجیرههای آب پیوسته در آوند چوبی که اغلب تحت کشش عظیم ایجاد شده توسط تعرق شکسته میشوند، مفید باشد.
نظریه های نیروی فیزیکی
این نظریه توسط دیکسون و جولی در سال ۱۸۹۴ ارائه شد و سپس توسط دیکسون در سال ۱۹۱۴ بهبود یافت. بنابراین، این نظریه به نام نظریه دیکسون در مورد صعود شیره نیز به نام او نامگذاری شده است. ویژگیهای اصلی نظریه عبارتند از:
پیوستگی ستون آب
ستون ممتد آب از ریشه از طریق ساقه و داخل برگها وجود دارد. ستون آب در عناصر آوندی وجود دارد. دومی به طور جداگانه عمل میکند اما یک سیستم پیوسته را از طریق مناطق بدون ضخامت خود تشکیل میدهد. از آنجایی که تعداد زیادی از عناصر آوندی در کنار هم قرار دارند، انسداد یک یا تعداد کمی از آنها باعث شکستگی در تداوم ستون آب نمیشود. ستون آب تحت تاثیر گرانش سقوط نمیکند زیرا نیروهای تعرق هم انرژی و هم کشش لازم را فراهم میکنند. پیوستگی، چسبندگی و کشش سطحی آب را در جای خود نگه میدارد.
چسبندگی یا مقاومت کششی
مولکولهای آب توسط یک نیروی جاذبه متقابل قوی به نام نیروی پیوستگی به یکدیگر متصل میمانند. جاذبه متقابل ناشی از پیوندهای هیدروژنی است که در بین مولکولهای آب مجاور ایجاد شده است. به دلیل نیروی پیوستگی، ستون آب میتواند تنش یا کشش تا ۱۰۰ اتمسفر را تحمل کند. بنابراین نیروی چسبندگی را استحکام کششی نیز مینامند. مقدار نظری آن حدود ۱۵۰۰۰ اتمسفر است اما مقدار اندازه گیری شده در داخل عناصر آوندی بین ۴۵ تا ۲۰۷ اتمسفر است. ستون آب به دلیل نیروی دیگری به نام نیروی چسبندگی بین دیوارههای بافتها و مولکولهای آب، اتصال خود را از عناصر آوندی (آوندها و تراکئیدها) قطع نمیکند. مولکولهای آب در حالت مایع بیشتر از مولکولهای آب در حالت گازی به یکدیگر جذب میشوند. این کشش سطحی ایجاد کرده که مویینگی بالایی را از طریق تراکئیدها و عناصر آوندی ایجاد میکند.
نیرویی کشش تعرق
فضاهای بین سلولی موجود در بین سلولهای مزوفیل برگها همیشه با بخار آب اشباع میشود. فضاهای بین سلولی مزوفیل از طریق روزنهها به هوای بیرون متصل میشوند. هوای بیرون به ندرت با بخار آب اشباع میشود و پتانسیل آب کمتری نسبت به هوای مرطوب موجود در داخل برگ دارد. بنابراین بخار آب از برگها ساطع میشود. سلولهای مزوفیل همچنان آب خود را به فضاهای بین سلولی از دست میدهند. در نتیجه کشش سطحی افزایش و پتانسیل آب (گاهی اوقات تا منفی ۳۰ بار) کاهش مییابد. سلولهای مزوفیل آب را از سلولهای عمیقتر خارج میکنند زیرا مولکولهای آن توسط پیوندهای هیدروژنی در کنار هم نگه داشته میشوند. سلولهای عمیق به نوبه خود از عناصر آوندی آب میگیرند. بنابراین، آب موجود در عناصر آوندی تحت کشش قرار میگیرد.
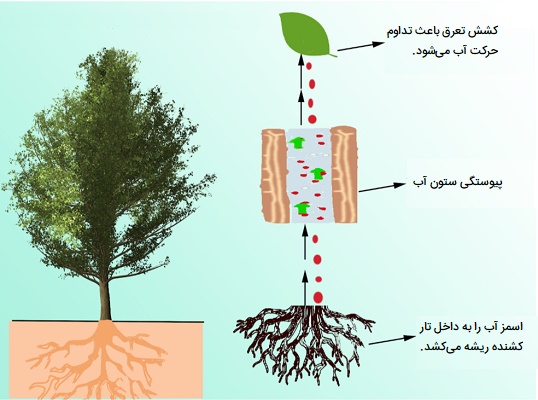
تنش مشابهی در میلیونها عنصر آوندی که در مجاورت سلولهای در حال تعرق قرار دارند احساس میشود که باعث میشود تمام ستون آب گیاه تحت کشش قرار گیرد. همانطور که کشش در اثر تعرق ایجاد میشود، به آن کشش تعرق نیز میگویند. با توجه به کشش ایجاد شده توسط تعرق، ستون آب گیاه به صورت منفعل از زیر به سمت بالای گیاه مانند طناب کشیده میشود.
شواهد تأیید کننده کشش تعرق
سرعت جذب آب و در نتیجه صعود شیره به شدت از سرعت تعرق پیروی میکند. ساقه متصل به لولهای که آب دارد و فرو رفتن در یک لیوان حاوی جیوه میتواند باعث حرکت جیوه به داخل لوله شود که کشش تعرق را نشان میدهد. در شاخهای که از یک گیاه سریع منتقل میشود، آب از انتهای بریده شده خارج میشود و نشان میدهد که ستون آب تحت کشش است. حداکثر کشش مشاهده شده در ستون آب ۱۰ تا ۲۰ اتمسفر است. این کشش کافی است که آب را به بالای بلندترین درختان حتی بیش از ۱۳۰ متر بکشد. تنش نمیتواند تداوم ستون آب را بشکند زیرا نیروی منسجم شیره آوند چوبی ۴۵ تا ۲۰۷ اتمسفر است. بازدانگان به دلیل وجود تراکئید به جای آوندها در صعود شیره خام در مضیقه قرار دارند. با این حال آوند چوبی ترکئید کمتر در معرض گرانش تحت کشش است. از این رو بیشتر درختان بلند دنیا را درختان سرخ و سوزنی برگ تشکیل میدهند.
پر بازدید ترین ها:
جزوه فوق العاده کابردی انترگال – از صفر تا صد تنها در ۳۰ صفحه